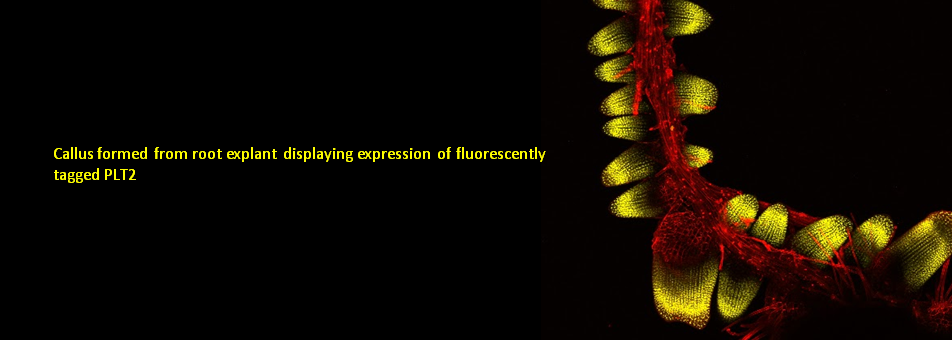
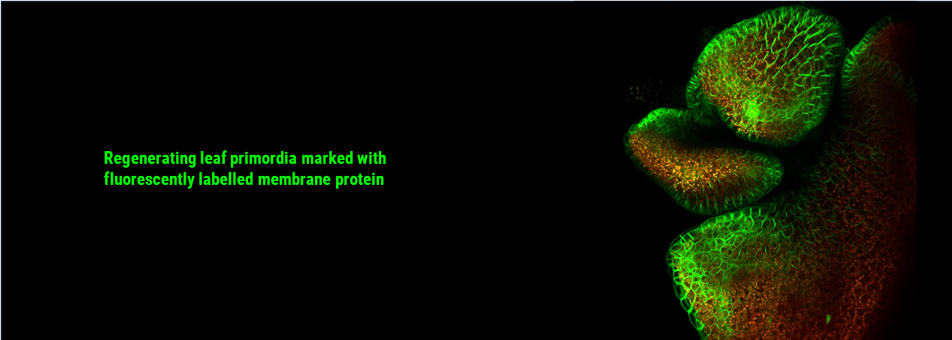
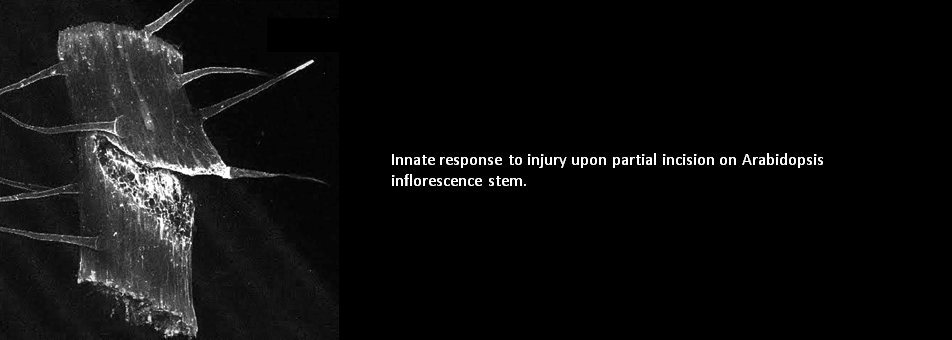
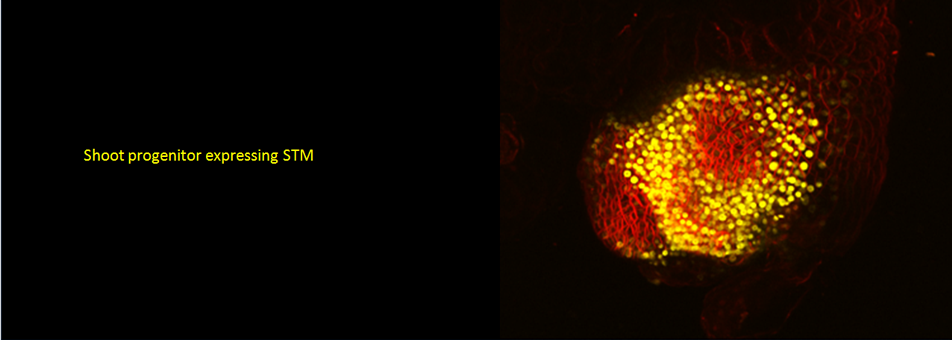
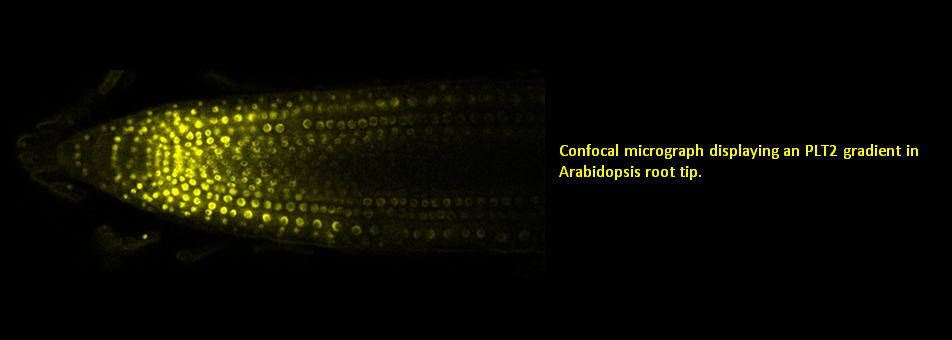
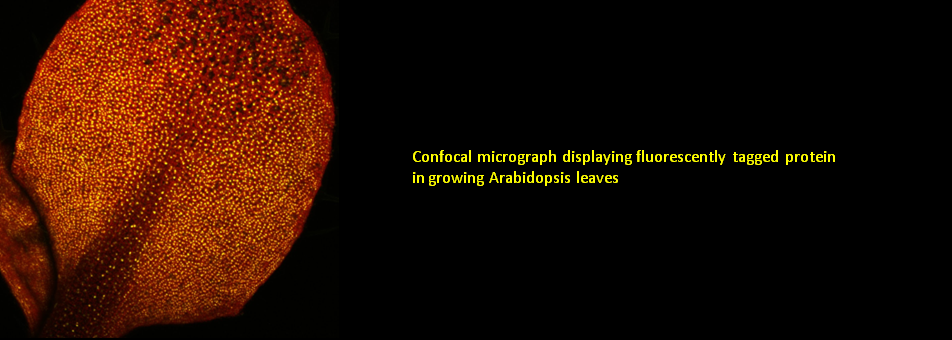
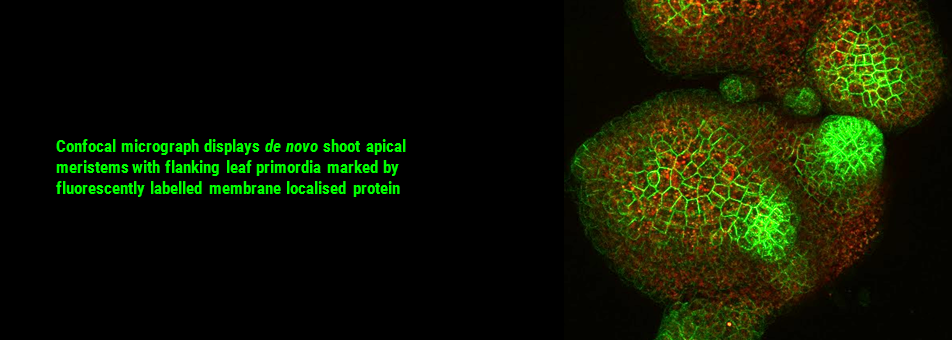
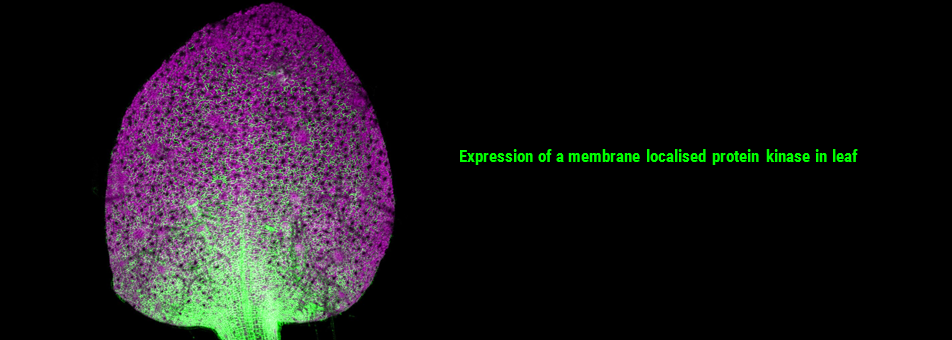
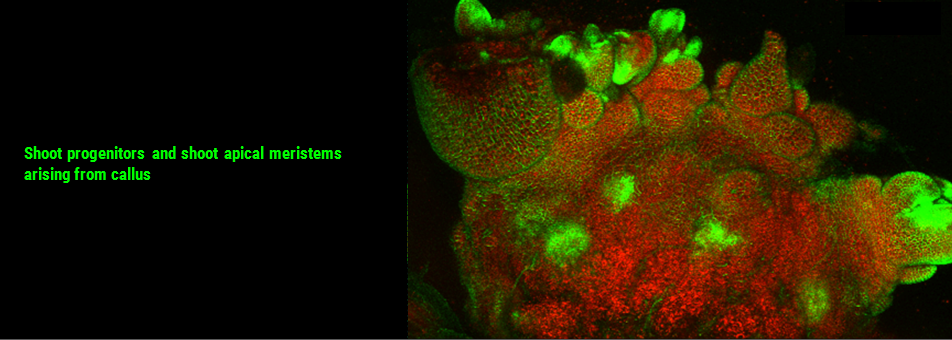
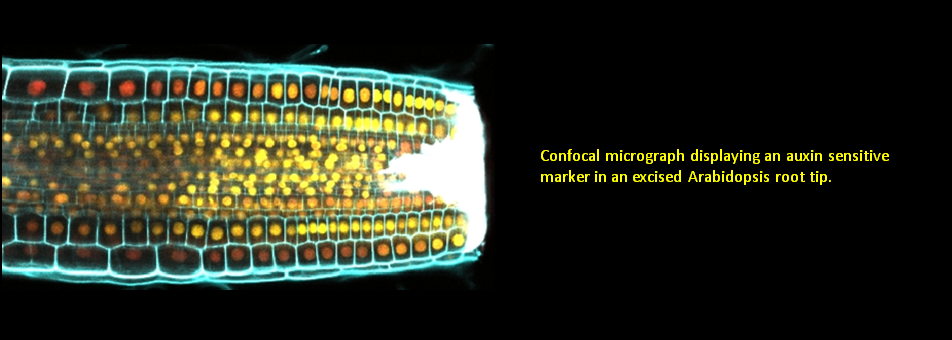
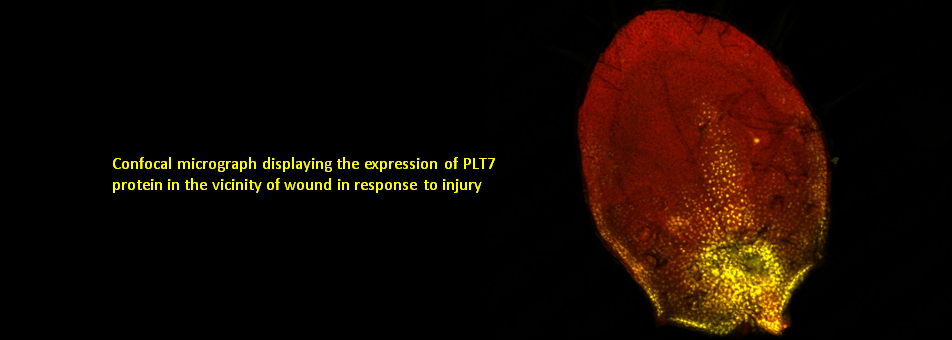
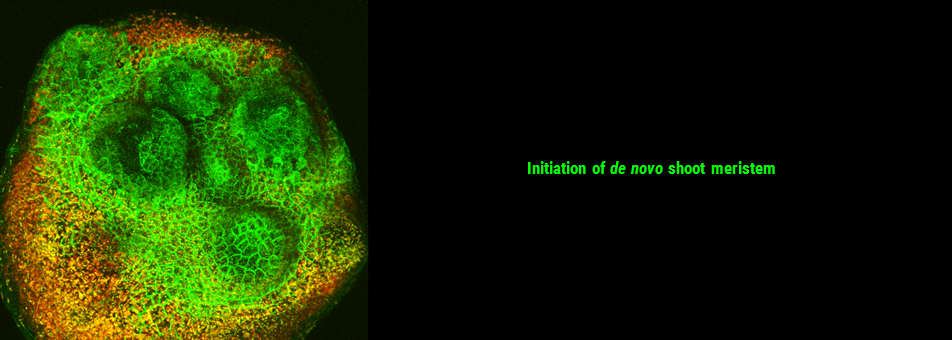
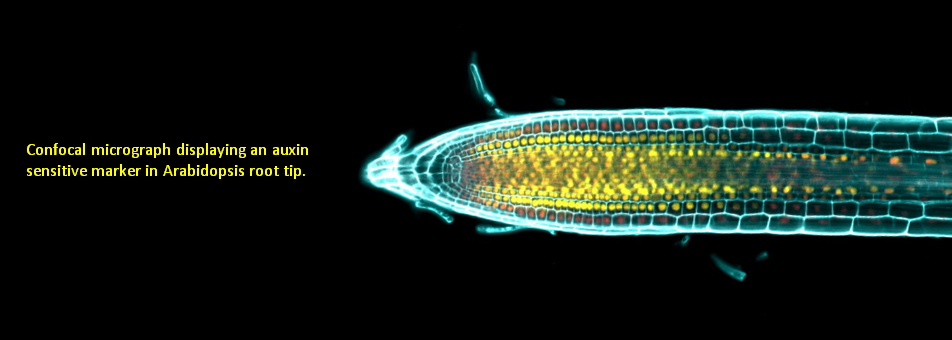
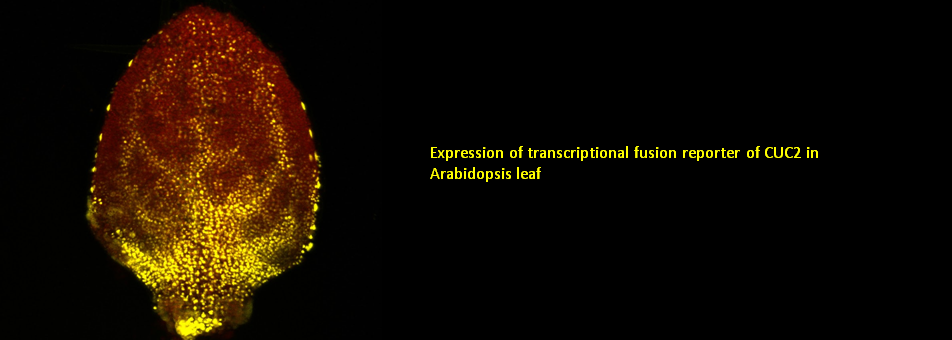
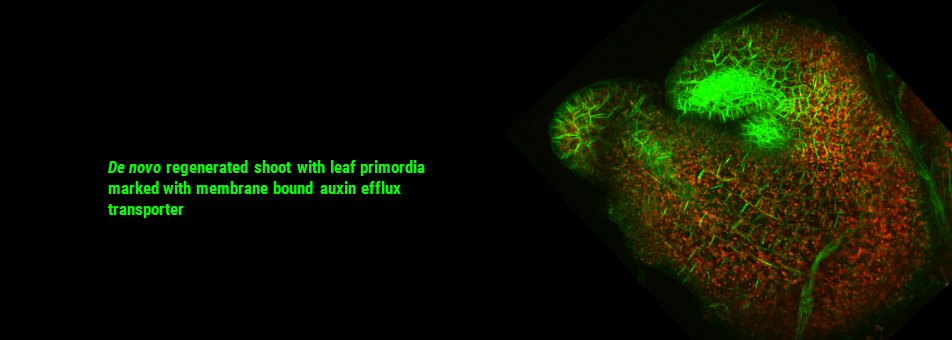
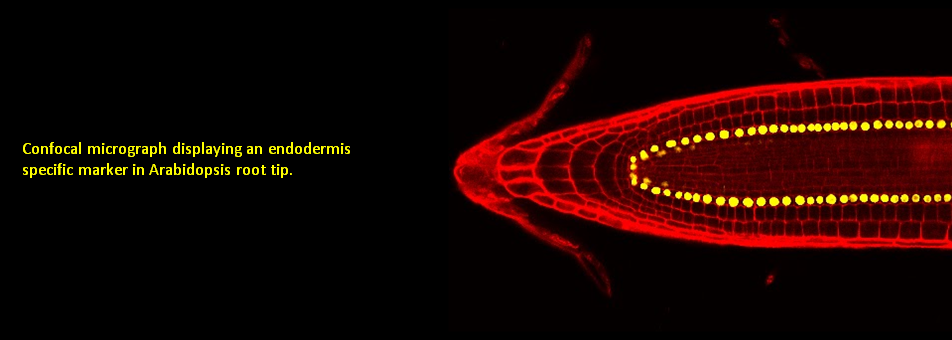
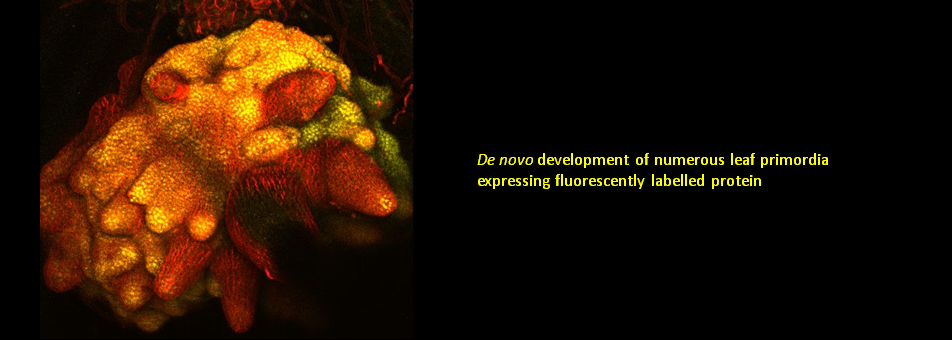
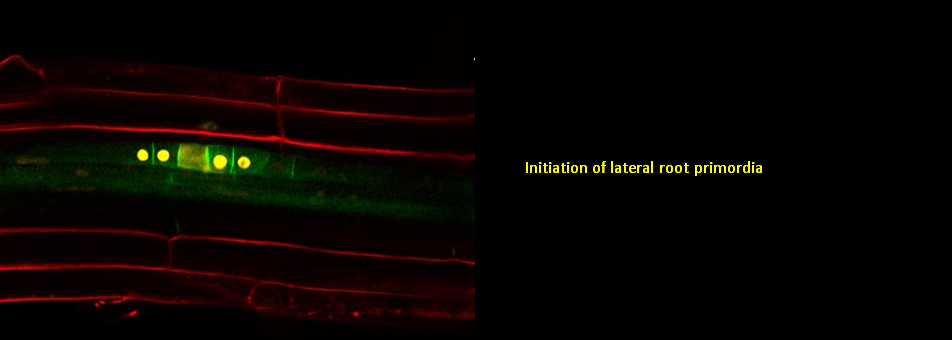
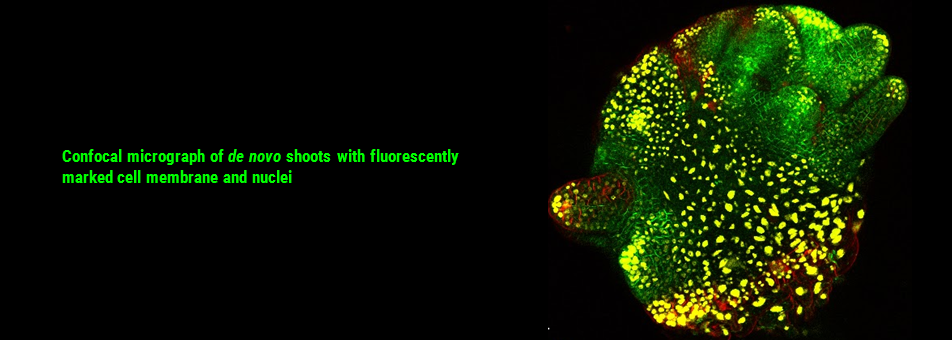
We use Arabidopsis as model to study plant regeneration. To learn more about this please read
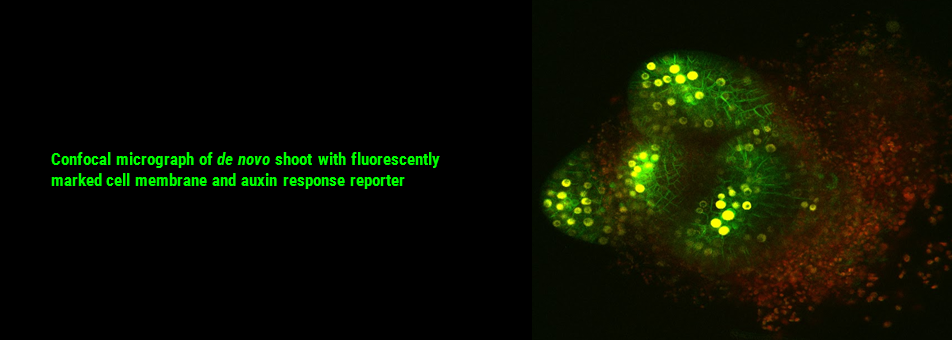
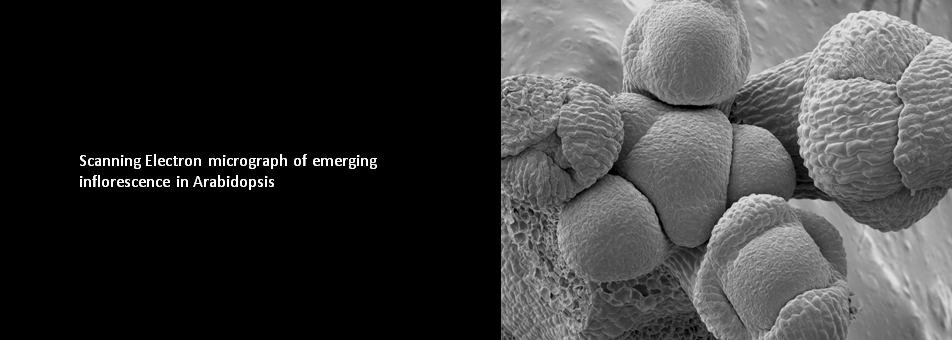
To learn more about de novo shoot regeneration please read following articles. • Mechanical conflict caused by a cell-wall-loosening enzyme activates de novo shoot regeneration. (https://www.sciencedirect.com/science/article/pii/S1534580722005482) • Shoot meristem progenitors emerge from mechanical heterogeneities. (https://doi.org/10.1016/j.devcel.2022.08.004)
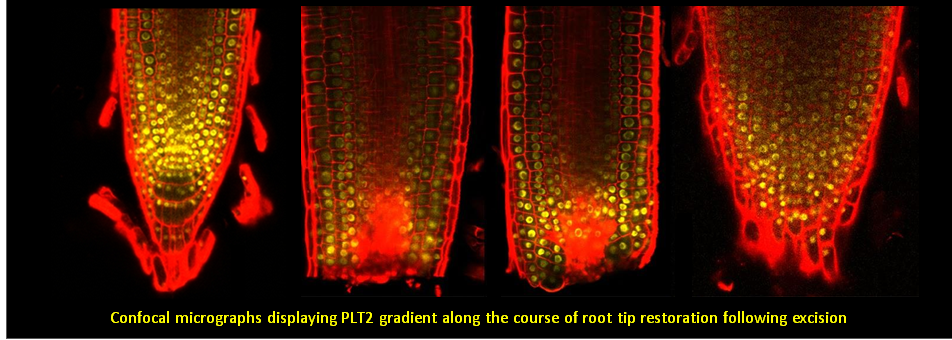
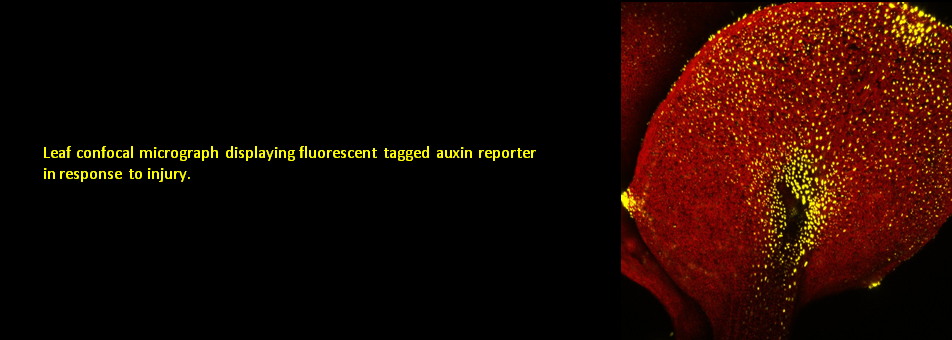
To learn more about regeneration of tissues or organs which were lost in injury, please read the following papers.
Want to learn more about our Research Highlights? click here
Recent Publications ● Ganguly, A., Humnabadkar, A., Gautam, K., Willemsen, V., Xu, L., Dagdas, Y., & Prasad, K*. (2026). PLETHORA–autophagy axis activates organ regeneration through ROS modulation. Proceedings of the National Academy of Sciences, 123(6), e2513954123. https://doi.org/10.1073/pnas.2513954123 (Featured on Cover, highlighted in Commentary: More than a blueprint: Developmental regulators secure the cellular environment for regeneration https://doi.org/10.1073/pnas.2600463123 (Xu J.,2026)) ● Mathew MM*, Saccheri J, Das S, Rajagopalan K, Lane B, Sidhardh KA, Smith RS, Willemsen V, Gomez MLG, Chakrabortty B, Scheres B, Tusscher KT*, Prasad K* (2025). Wound repair in plants guided by cell geometry. Current Biology, 3851-3868.e7. https://doi.org/10.1016/j.cub.2025.06.072 Highlights: https://www.nature.com/articles/d44151-025-00167-2 ) ● Varapparambath V, Mathew MM, Shanmukhan AP, Radhakrishnan D, Kareem A, Verma S, Ramalho JJ, Manoj B, Vellandath AR, Aiyaz M, Radha RK, Landge AN, Mähönen AP, Heisler MG, Weijers D, Prasad K*. (2022). Mechanical conflict caused by a cell-wall-loosening enzyme activates de novo shoot regeneration. Developmental Cell. 12;57(17):2063-2080.e10. https://doi.org/10.1016/j.devcel.2022.07.017 (Featured on Cover, Featured in Journal, highlighted in Preview: Shoot meristem progenitors emerge from mechanical heterogeneities https://doi.org /10.1016/ j.devcel. 2022.08.004 (Hamant, 2022), highlighted in Faculty Opinions: https:// facultyopinions.com/ article/ 74229178)
Vacancies Inquiries from prospective students, graduate trainees, postdocs and visitors are welcome. Please email on kalika.prasad@iiserpune.ac.in with your detailed cv and an outline of your interests.